Energía, oxígeno y forma: una lectura evolutiva del sistema respiratorio terrestre y sus posibles variantes biológicas fuera de la Tierra
Introducción
La historia del sistema respiratorio suele contarse desde el pulmón. Es comprensible: el pulmón representa, en los mamíferos, la culminación anatómica más familiar del intercambio gaseoso. Sin embargo, esa manera de narrar el problema invierte el orden biológico real. La respiración no comenzó con órganos especializados ni con aire rico en oxígeno. Comenzó con una pregunta más elemental: cómo sostener la vida cuando la energía disponible es limitada y el entorno impone restricciones fisicoquímicas concretas. En ese sentido, el sistema respiratorio no debe entenderse como un punto de partida, sino como una solución tardía dentro de una historia mucho más extensa, donde primero emergen la bioquímica, luego la organización celular, después la multicelularidad y, solo entonces, las estructuras anatómicas dedicadas al transporte y al intercambio de gases.
La Tierra primitiva ofrece el marco más instructivo para entender este proceso. La vida apareció y persistió durante largos intervalos sin depender del oxígeno libre atmosférico. Más aún, la acumulación de oxígeno fue en un inicio un problema antes que una ventaja. Solo tras una larga transición geoquímica y biológica, el oxígeno pudo convertirse en el soporte de una bioenergética más eficiente y, con ello, en uno de los prerrequisitos de la complejidad multicelular. El paso desde metabolismos anaerobios hacia sistemas oxidativos de alto rendimiento no solo cambió el rendimiento energético de la vida; rediseñó la arquitectura de los organismos y, con ella, la necesidad de sistemas especializados de intercambio gaseoso. La revisión de Lindahl sobre oxígeno y vida en la Tierra lo resume bien: el oxígeno libre aumentó progresivamente en la atmósfera durante la historia geológica y, a la vez, la biología tuvo que desarrollar mecanismos para sensarlo, utilizarlo y contener su reactividad. (PubMed)
Desde esa perspectiva, estudiar la evolución del sistema respiratorio obliga a unir niveles que con frecuencia se abordan por separado: geoquímica planetaria, metabolismo celular, endosimbiosis, multicelularidad, anatomía comparada y desarrollo embrionario. También permite abrir, con algo más de rigor, la vieja pregunta sobre cómo podría haberse resuelto este mismo problema en otros mundos. Si la complejidad biológica surgiera fuera de la Tierra, no tendría por qué copiar nuestros órganos; pero difícilmente escaparía a la necesidad de encontrar un equilibrio viable entre disponibilidad energética, gradientes gaseosos, presión ambiental, gravedad y ontogénesis. Ese será el hilo conductor del presente ensayo.
Orígenes: vida sin oxígeno y metabolismo bajo restricciones
Los primeros sistemas vivos surgieron en un planeta cuya atmósfera era muy distinta de la actual. La evidencia geológica y biológica indica que la vida apareció antes de la acumulación sustancial de oxígeno libre en la atmósfera y que durante una parte muy prolongada de la historia terrestre los metabolismos dominantes fueron anaerobios. La Gran Oxidación suele situarse alrededor de 2.45–2.32 mil millones de años atrás, muy posterior al origen de la vida y probablemente posterior también al origen de linajes cianobacterianos capaces de fotosíntesis oxigénica. (PMC)
Esto tiene una consecuencia conceptual importante. La vida no necesitó oxígeno para comenzar. Lo que sí hizo falta fue una bioquímica capaz de capturar energía del entorno y mantener organización interna suficiente para sostener replicación, reparación y evolución. En términos respiratorios estrictos, eso significa que el intercambio gaseoso especializado no fue necesario al inicio porque la escala del problema aún era pequeña: organismos unicelulares o comunidades simples podían resolver sus gradientes por difusión y mediante rutas metabólicas de rendimiento modesto.
Pero esos metabolismos tenían un techo. La vida anaerobia puede ser extraordinariamente resistente, incluso hoy; los extremófilos lo muestran de manera contundente. Sin embargo, la supervivencia y la complejidad no son sinónimos. El problema no radicaba en la posibilidad de existir, sino en la dificultad de escalar. Sin una bioenergética de mayor rendimiento, la vida podía persistir, diversificarse y adaptarse, pero no sostener con facilidad tejidos extensos, integración funcional compleja o altos costos de mantenimiento.
La Gran Oxidación y la paradoja del oxígeno
La acumulación de oxígeno libre representó un cambio de régimen planetario. Fue el producto de la fotosíntesis oxigénica y no una condición inicial del ambiente. La literatura sobre cianobacterias y Gran Oxidación ha mostrado no solo que estos linajes anteceden al evento, sino también que la multicelularidad cianobacteriana y la diversificación morfológica guardan relación temporal con el ascenso del oxígeno atmosférico. (PMC)
Desde el punto de vista biológico, el oxígeno impuso una paradoja que sigue vigente hasta hoy. Permitió una extracción energética mucho mayor por unidad de sustrato, pero al mismo tiempo introdujo una amenaza oxidativa permanente. La vida tuvo que desarrollar mecanismos de defensa, sistemas antioxidantes, regulación fina del uso de oxígeno y estrategias de compartimentalización. El oxígeno no fue adoptado como un “bien puro”; fue integrado como una molécula útil y peligrosa a la vez. Ese doble carácter sigue siendo visible en la fisiología respiratoria contemporánea, en la que tanto la hipoxia como la hiperoxia son dañinas y el valor biológico del oxígeno depende siempre de contexto, concentración, tiempo de exposición y capacidad tisular para manejarlo. (PubMed)
La trascendencia de esta transición no radica solo en la atmósfera. Radica en que, una vez disponible el oxígeno, la vida pudo apoyarse en una nueva economía energética.
La mitocondria: internalización de la alta energía
El salto decisivo no fue simplemente “tener oxígeno alrededor”, sino internalizar de manera eficaz la fosforilación oxidativa. Ese es el papel histórico de la mitocondria. La endosimbiosis que dio origen a las mitocondrias permitió que células eucariotas accedieran a una forma mucho más eficiente de producción de ATP. Aunque la complejidad de la transición eucariota excede este ensayo, el punto fisiológico es simple: con mitocondrias, la célula dejó de depender exclusivamente de una economía energética restringida y pudo sostener mayor tamaño, más organización interna y funciones más costosas.
Ese cambio alteró profundamente el horizonte evolutivo. La multicelularidad dejó de ser una rareza limitada y se volvió una posibilidad estable. La revisión comparativa sobre evolución de la respiración aérea lo expresa de manera indirecta al mostrar que la historia de los órganos respiratorios es inseparable de la historia previa de la organización corporal y de las demandas metabólicas que esa organización impone. (PMC)
La secuencia, por tanto, no fue: aparece el pulmón y luego la complejidad. Fue al revés: la biología adquiere una base energética suficiente, crece en escala y entonces necesita órganos de intercambio.
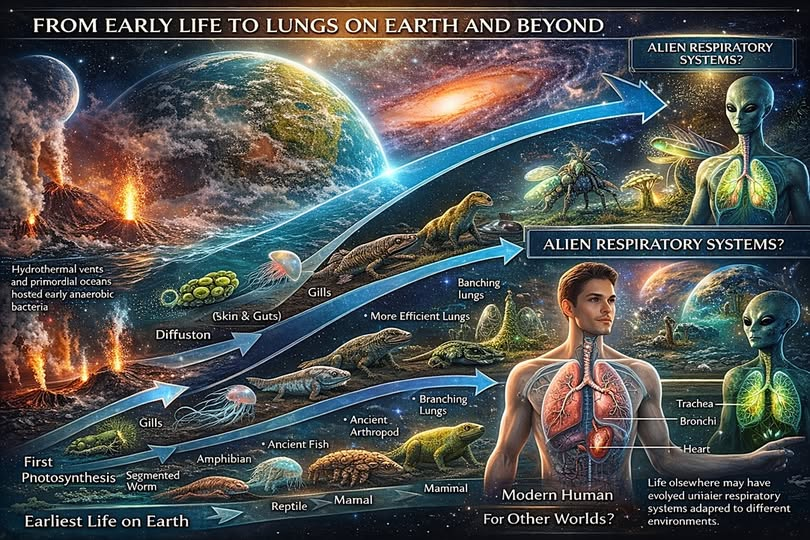
De la difusión a los órganos: cuando la geometría deja de ser suficiente
En organismos pequeños, la difusión puede bastar. A medida que aumenta la distancia entre el medio externo y las células internas, la difusión se vuelve insuficiente. Ahí comienza realmente la historia anatómica del sistema respiratorio. La evolución terrestre produjo múltiples soluciones, no una sola, y cada una refleja una combinación particular de medio, tamaño corporal, demanda metabólica y restricciones mecánicas.
En peces y otros organismos acuáticos, las branquias resolvieron el problema mediante superficies amplias, delgadas y altamente perfundidas, capaces de aprovechar el oxígeno disuelto en un medio mucho más denso que el aire, pero también mucho menos rico en oxígeno utilizable. En ese contexto, el intercambio exige gran área, flujo continuo y un diseño expuesto pero eficaz. La transición hacia respiración aérea en vertebrados no ocurrió de manera lineal ni uniforme; los estudios sobre evolución del pulmón en vertebrados muestran trayectorias complejas y no una única innovación repentina. (PMC)
En artrópodos terrestres, especialmente insectos, la solución fue radicalmente distinta. No se desarrolló un pulmón vascularizado al estilo vertebrado, sino un sistema traqueal que lleva el gas casi directamente a los tejidos. La revisión de Maina y West sobre la evolución de la respiración aérea enfatiza que los invertebrados desarrollaron una gran variedad de órganos: tráqueas, pulmones en libro, respiración cutánea, branquias o combinaciones de ellos, siempre ajustados a las condiciones específicas de cada linaje. (PMC)
En aves, el sistema respiratorio representa una de las arquitecturas más eficientes conocidas. El flujo unidireccional, acoplado a sacos aéreos, mantiene gradientes favorables durante todo el ciclo ventilatorio. No es casual que esta solución aparezca en organismos con alta demanda energética sostenida. El sistema aviar no es una versión “mejor” del pulmón mamífero en términos absolutos; es una respuesta específica a una ecología y a una economía energética concretas. (PMC)
Los mamíferos, finalmente, desarrollaron pulmones alveolares ventilados por un patrón tidal. Su eficiencia teórica es menor que la del flujo unidireccional aviar, pero ofrecen versatilidad mecánica, compatibilidad con un cuerpo altamente móvil y una integración funcional adecuada con la circulación y el metabolismo mamíferos. El punto importante aquí no es decidir qué órgano es superior, sino reconocer que cada solución nace de una biología previa. La forma del sistema respiratorio nunca es independiente del organismo que lo porta.
El caso humano: del ambiente hipóxico temprano al pulmón adulto
La embriología humana añade una dimensión esencial a esta historia. En el ser humano, el desarrollo inicial no ocurre en un ambiente de alta oxigenación. Durante la invasión uterina temprana, el trofoblasto extravelloso forma tapones que limitan la entrada de sangre materna al espacio intervelloso, manteniendo un ambiente de baja tensión de oxígeno durante la organogénesis. Las mediciones clásicas de Jauniaux y colaboradores mostraron tensiones de oxígeno placentarias bajas en el primer trimestre, con incremento posterior cuando la circulación materna se establece de forma más plena hacia las semanas 12–13. Ese ambiente relativamente hipóxico no es una patología; participa en la regulación del desarrollo embrionario temprano. (PMC)
Esto corrige una simplificación frecuente. El desarrollo humano no está programado para un máximo temprano de oxígeno. Está programado para un manejo secuencial del oxígeno. El hecho de que la organogénesis humana transcurra bajo tensiones bajas recuerda que la biología no busca “más oxígeno” en abstracto, sino el gradiente adecuado para cada etapa. El sistema respiratorio humano, visto desde esta óptica, no debe definirse solo por el pulmón adulto y su intercambio alveolar, sino por toda la trayectoria de desarrollo que conduce hasta él.
El pulmón adulto humano es, por tanto, la fase final de una solución mamífera terrestre a un problema energético y geométrico de gran escala. Requiere diafragma, caja torácica, circulación pulmonar de alta capacidad, hemoglobina con afinidad ajustada, control central de la ventilación y una interfaz alveolocapilar extremadamente delgada. Pero ninguna de esas piezas tendría sentido si antes no hubieran existido una bioquímica oxidativa eficiente, una ontogénesis regulada por gradientes apropiados y un ambiente atmosférico estable en torno a 21% de oxígeno.
De la Tierra al cosmos: cómo pensar un sistema respiratorio en otros mundos
Una vez entendida la historia terrestre, la extrapolación hacia otros mundos se vuelve más rigurosa. La cuestión no es imaginar “pulmones alienígenas” como simples variantes morfológicas de los nuestros. Lo que debe imaginarse primero es la biología que haría necesarios esos órganos. Stratmann lo plantea bien desde otro ángulo: cualquier extrapolación útil en ciencia ficción médica tiene que partir de cómo funciona el cuerpo real, de sus límites y de las condiciones que lo moldean.
Bajo esa lógica, un sistema respiratorio extraterrestre dependería de al menos nueve variables mayores.
La primera sería la fuente y disponibilidad energética. Sin un salto bioenergético equivalente al que en la Tierra representó el uso controlado del oxígeno, la vida compleja e inteligente sería menos probable. Podría haber vida microbiana o colonial sin un sistema respiratorio elaborado, pero sostener gran tamaño, movilidad continua o procesamiento neural avanzado exigiría una química energética de alto rendimiento.
La segunda sería la gravedad. En baja gravedad, la ventilación podría requerir menos trabajo mecánico y favorecer estructuras finas y extensas; en alta gravedad, las cargas sobre pared torácica, circulación y distribución interna del gas exigirían sistemas más compactos, quizá más rígidos o con soporte estructural mayor. La experiencia humana en microgravedad muestra que el pulmón tolera bien la ausencia de gravedad, pero también confirma que el entorno modifica distribución de fluidos, ventilación y eliminación de CO₂, lo que sugiere que una especie evolucionada permanentemente en otra gravedad seguiría una trayectoria distinta. (PMC)
La tercera sería la presión atmosférica total. La biología responde a presiones parciales, no solo a porcentajes. Un planeta con baja fracción de oxígeno, pero atmósfera densa podría ofrecer una pO₂ suficiente; otro con alta fracción de oxígeno, pero atmósfera tenue podría seguir siendo restrictivo. Ese principio, tan familiar en fisiología de altura, sería universal.
La cuarta sería la composición química del medio. El oxígeno no es la única posibilidad teórica, aunque para vida compleja sigue siendo la opción más eficaz conocida. Si otra especie usara otro aceptor final de electrones, cambiarían la bioquímica del intercambio, los pigmentos transportadores, el manejo ácido-base y la toxicidad ambiental. La ciencia ficción ha imaginado especies que respiran cloro o viven en medios basados en otras químicas, pero el valor real de esas imágenes no está en el exotismo, sino en recordar que cambiar el gas implica rehacer toda la fisiología.
La quinta sería la densidad del medio externo. Respirar en un gas ligero no plantea el mismo problema que hacerlo en una atmósfera muy densa o en un medio líquido. A mayor densidad, aumentan algunas resistencias, pero también puede cambiar la estabilidad de superficies de intercambio. Los órganos respiratorios no existen aislados de la mecánica de fluidos.
La sexta sería la demanda metabólica del organismo. Una especie de metabolismo lento podría sostenerse con gradientes modestos y gran tolerancia a hipoxia relativa. Una especie muy activa necesitaría intercambio rápido, transporte eficiente y control estricto del gas y del calor.
La séptima sería la escala corporal. La difusión sirve para organismos pequeños; deja de servir en organismos grandes si no aparece una solución anatómica o circulatoria. Esta relación entre tamaño y necesidad de órganos de intercambio seguiría siendo válida fuera de la Tierra.
La octava sería la protección de la interfaz respiratoria. Toda superficie que intercambia gases es vulnerable a desecación, infección, oxidación, colapso o daño físico. El sistema respiratorio extraterrestre tendría que resolver ese costo, como lo hace el nuestro con moco, barreras epiteliales, surfactante, defensa inmune y regulación del flujo.
La novena, y quizá menos considerada, sería la ontogénesis. El ejemplo humano muestra que el desarrollo inicial puede requerir tensiones de oxígeno bajas y reguladas. Una especie alienígena podría madurar en un microambiente distinto al del adulto, con etapas de hipoxia programada o con exposición secuencial a gases diferentes. Su sistema respiratorio no debería imaginarse solo desde el adulto funcional, sino desde el proceso por el que se construye.
Discusión
Vistas en conjunto, estas consideraciones llevan a una conclusión sobria. La Tierra no demuestra que el pulmón mamífero sea la forma inevitable de la respiración compleja. Demuestra otra cosa: que la complejidad biológica necesita resolver el problema energético y luego el problema del intercambio. La secuencia importa. Primero se consolida una bioquímica capaz de sostener complejidad; luego aparece la anatomía que la vuelve viable a escala del organismo completo.
En otros mundos, si la vida compleja surgiera, sería razonable esperar el mismo orden general, aunque no las mismas estructuras. Podrían existir sistemas de flujo unidireccional, redes traqueales profundas, interfaces distribuidas, membranas extensas sostenidas por fluidos más densos o combinaciones hoy desconocidas. Lo que difícilmente cambiaría es la necesidad de coordinar disponibilidad energética, gradientes de gas, protección tisular y desarrollo.
Esta forma de pensar no solo ordena la especulación. También devuelve cierta humildad a la fisiología terrestre. Nuestro sistema respiratorio no es “el” sistema respiratorio; es una de las respuestas posibles, moldeada por 1 g, por una atmósfera concreta, por una historia de oxigenación planetaria particular y por una embriología que comienza en relativa hipoxia y culmina en un pulmón alveolar altamente oxidativo.
Conclusión
La evolución del sistema respiratorio en la Tierra puede leerse como la historia de cómo la vida aprendió a manejar una nueva forma de energía sin ser destruida por ella. Los extremófilos y los primeros anaerobios muestran que la vida puede comenzar con poco. La Gran Oxidación muestra que el entorno puede abrir posibilidades bioenergéticas inéditas. La mitocondria muestra que la energía de alta densidad debe ser internalizada y regulada. Las branquias, tráqueas, pulmones aviares y pulmones mamíferos muestran que no existe una única solución anatómica, sino respuestas distintas a presiones distintas. La embriología humana recuerda que el oxígeno no solo se usa: también se administra en el tiempo del desarrollo.
Si en otras regiones del universo hubiera surgido vida inteligente, su sistema respiratorio no tendría por qué parecerse al nuestro. Pero sí tendría que ser la consecuencia de la misma clase de problema: cómo sostener organización compleja cuando la energía deja de ser marginal y el entorno exige un intercambio gaseoso compatible con la supervivencia, el crecimiento y la reproducción. Ahí radica, probablemente, el verdadero universal biológico.
Referencias
Lindahl SGE. Oxygen and life on earth: an anesthesiologist’s views on oxygen evolution, discovery, sensing, and utilization. Anesthesiology. 2008;109:7–13. (PubMed)
Semenza GL. Life with oxygen. Science. 2007;316:62–64.
Jauniaux E, Watson AL, Hempstock J, Bao YP, Skepper JN, Burton GJ. Onset of maternal arterial blood flow and placental oxidative stress. Am J Pathol. 2000;157:2111–2122. (PMC)
Schirrmeister BE, de Vos JM, Antonelli A, Bagheri HC. Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event. Proc Natl Acad Sci U S A. 2013;110:1791–1796. (PubMed)
Maina JN, West JB. Evolution of air breathing: oxygen homeostasis and the transitions from water to land and sky. Compr Physiol. 2013;4:849–915. (PMC)
Hoffman M, Taylor BE, Harris MB. Evolution of lung breathing from a lungless primitive vertebrate. Respir Physiol Neurobiol. 2016;224:11–16. (PMC)
Ortiz-Prado E, Dunn JF, Vasconez J, Castillo D, Viscor G. Partial pressure of oxygen in the human body: a general review. Am J Blood Res. 2019;9:1–14.
Stratmann HG. Using Medicine in Science Fiction: The SF Writer’s Guide to Human Biology. Cham: Springer; 2016.